Reciprocal interactions between malignant and stromal cells create a local microenvironment that fosters tumor growth. Extracellular vesicles (EVs) such as exosomes, microvesicles, and large oncosomes are involved in tumour-stroma communication by shuttling signaling cargo and other molecules. EVs take part in the response to anticancer therapeutics not least by promoting drug resistance throughout the targeted tumor. Moreover, circulating EVs can also transport important molecules to remote destinations in order to prime metastatic niches in an otherwise healthy tissue.
- STEP 1: Isolation and characterisation of nanoparticles/extracellular vesicles by Nanosight and Exoview R100
-
Rapid, automated analysis of the size distribution and concentration of all types of nanoparticles (10nm to 2000nm) using the Nanosight technology.
We provide particle analysis data (in triplicate) in reports that show the mean, mode diameter size and concentration of nanoparticles. In addition, during data collection, a video is recorded from each independent sample (also supplied as wmv file).

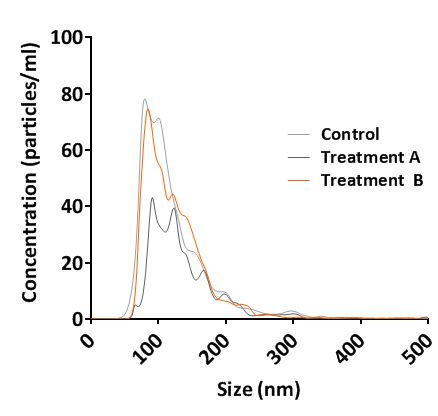
Nanosight NS300 Analysis Report Laser light scattering tracking
of exosomes
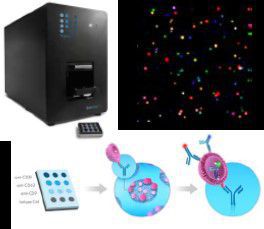
Exoview detection of biomarkers on single EVs. Capture-antibody-functionalized microarray chips and fluorescent antibodies can be used for transmembrane and/or cargo labelling of specific proteins (biomarkers).
- STEP2: Visualisation and validation of exosomes
-
Exosomes are isolated by ultracentrifugation or by using the ExoQuick (serum, plasma, ascites samples) or ExoQuick-TC (cell media, urine, spinal fluid) kits.
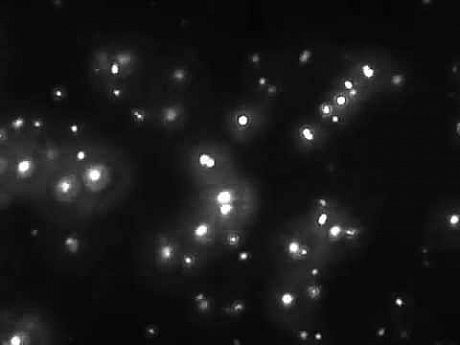
Validation of exosomes isolation can be provided via: i) Western blotting for selected markers (i.e. CD9, CD63, CD81), ii) Transmission Electron Microcrospy visualisation and iii) Immunofluorescence microscopy following PKH26 labelling of isolated exosomes.
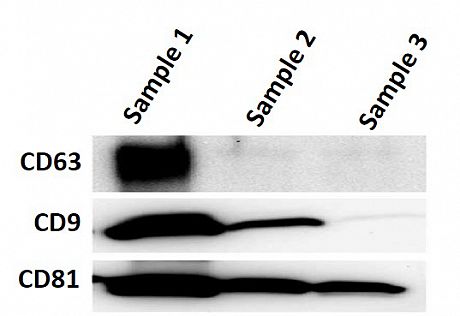
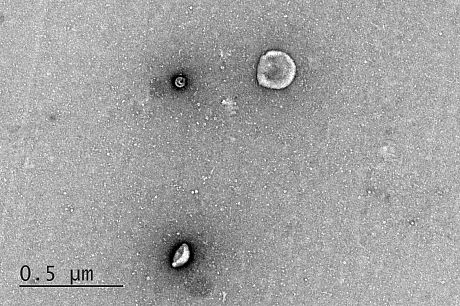
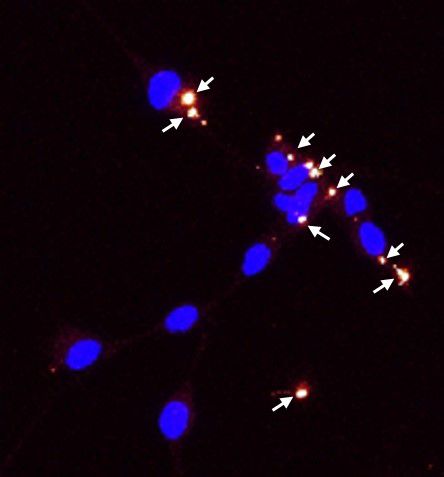
WB of exosomal protein markers Exosomes analysis by TEM PKH26 labelled exosomes
- STEP3: Identification of proteomic content of exosomes by mass spectrometry and Data analysis
-
Identification of various exosome surface and internal proteins will be assessed by mass spectrometry analysis. Data are analysed using MASCOT databases and Scaffold software. Additional bioinformatics analysis / comparison of identified proteins can be provided.

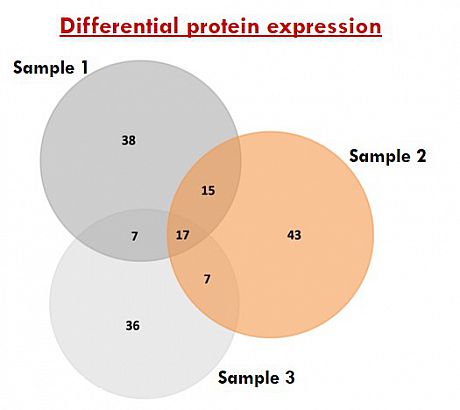
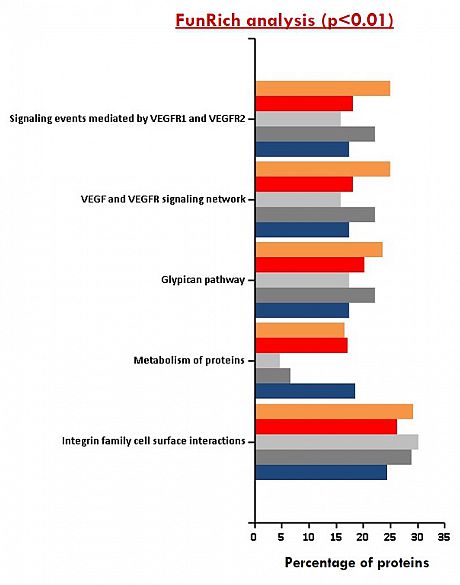
Table of exosome proteins MS/MS data Venn diagram distribution of common Functional enrichment analysis and unique exosomal proteins of exosomal proteins
- PAPER: Shedding of bevacizumab in tumour cells-derived extracellular vesicles as a new therapeutic escape mechanism in Glioblastoma (Molecular Cancer, 2018)
-
GBM is amongst the most aggressive types of brain tumours for which current treatments are of limited benefit. During the past decades, AAT have provided a rationale for targeting and blocking the tumour blood supply. Unfortunately, the effects of AAT/bevacizumab, a monoclonal humanised antibody neutralising Vascular Endothelial Growth Factor-A (VEGF-A), on tumour growth are short-term and GBM patients ultimately relapse. Interestingly, since the expression of some pro-angiogenic factors and their receptors (i.e. VEGF-A/VEGF-R) has been described in tumour cells, it appears that AAT/bevacizumab also acts directly on GBM cells that might eventually lead to therapy resistance and relapse.
Here, we demonstrated that Bevacizumab affects the EVs proteomic content derived from GBM cells (Fig 1).Moreover, Bevacizumab can be internalised by GBM cells (Fig 2), while it is also detectable at the surface of GBM cells-derived EVs (Fig 3).
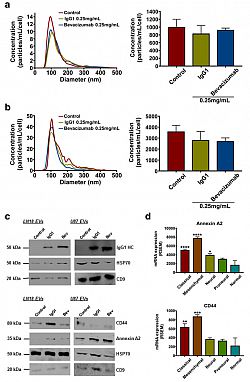
Fig. 1 IgG1/Bevacizumab antibody can affect LN18 and U87 GBM cells-derived EVs concentration and their proteomic content. NTA of (a) LN18 or (b) U87 GBM cells-derived EVs following treatment with bevacizumab (0.25 mg/mL). LN18 or U87 GBM cells were treated for 24 h with 0.25 mg/ mL IgG1 or bevacizumab. Then cells were washed two times with sterile PBS and incubated additional 24 h in serum free conditions without treatment. CM was then collected, EVs were isolated and re-suspended in 100 μL filtered sterile PBS. EVs suspension was 1/5 diluted and infused to a Nanosight© NS300 instrument. 5 captures of 60s each were recorded. Particles concentration (particles/mL) and size (nm) were measured. Particles concentration was normalised to the number of cells after treatment (particles/mL/cell). The mean ± SEM of 3 independent experiments is shown. (c) Western blotting validation of the human IgG, Annexin A2 and CD44 in EVs derived from LN18 and U87 GBM cells. (d) Gene expression distribution of Annexin A2 and CD44 among the different GBM subtypes has been obtained from TCGA. The mean ± SEM is shown (*p < 0.05, **p < 0.01, ***p < 0.001,****p < 0.0001; ANOVA, compare to ‘normal’).
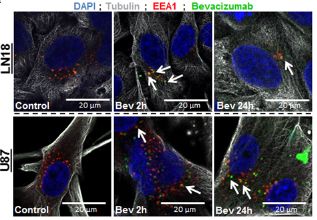
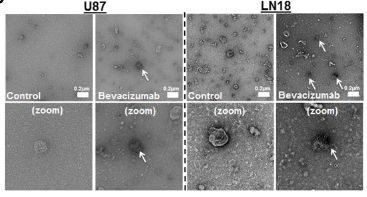
Fig. 2 (upper left) Bevacizumab is internalised by GBM cells Immunofluorescence detection of bevacizumab and EEA1 in LN18 and U87 GBM cells. GBM cells were allowed to grow on cover slips and then treated with 0.25 mg/mL bevacizumab for 2 h and 24 h. Cells were fixed with 4% PFA and then incubated with antibodies against α-tubulin, EEA1 and human IgG1.
Fig. 3 (upper right) Bevacizumab is detectable at the surface of GBM cells-derived EVs following treatment TEM detection of bevacizumab in LN18 and U87 GBM cells-derived EVs. U87 GBM cells were treated for 24 h with 0.25 mg/mL bevacizumab. Then cells were washed two times with sterile PBS and incubated additional 24 h in serum free conditions without treatment. CM was then collected and EVs were isolated. Immuno-gold labeling was then performed against human IgG in the EVs fractions.
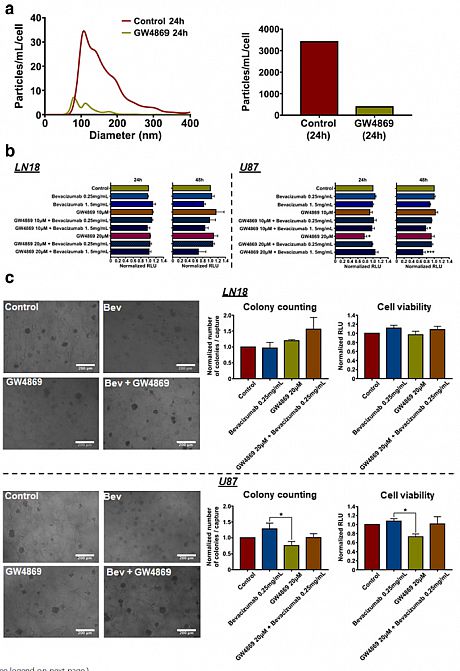
Moreover, we provide evidence showing that inhibition of EVs production increases the effects of bevacizumab on GBM cells’ viability (Fig 4). Taken together, we report that the paradoxical pro-invasive effect of bevacizumab on GBM cells might be due to alterations in the tumour cells-derived EVs, including shedding of the antibody and further modifications of the cargo, both possibly contributing to therapeutic resistance. Therefore, this study suggests the combination of bevacizumab with a local blocking of EVs-dependent intercellular communication as a potential new therapeutic strategy to improve GBM treatment.
Fig. 4 Inhibition of EVs production increases effects of bevacizumab on U87 GBM cells' viability. (a) NTA of LN18 GBM cells-derived EVs following treatment with GW4869 (20 μM). Particles concentration was normalised to the number of cells after treatment (particles/mL/cell). (b) LN18 and U87 GBM cells' viability assay in response to bevacizumab combined with GW4869. Results are expressed as normalised Relative Light Unit (RLU). (*p < 0.05, ***p < 0.01; ANOVA). (c) LN18 and U87 GBM cells' invasiveness assay using a hyaluronic acid (HA) hydrogel. Results are expressed as normalised number of colonies / capture and normalised RLU, respectively. (*p < 0.05; ANOVA)